狗的衰老:分子生物学机器翻译
宠物狗衰老过程中的血液生物标志物与品种遗传学
Vista Sohrab¹,²,³*, Michelle E. White³, Benjamin R. Harrison⁴, Rob Bierman⁵, Abbey Marye⁶, Kathleen Morrill Pirovich¹,²,³,⁷, Diane P. Genereux³, Kate Megquier³, Xue Li¹,²,³,⁸, Brittney Kenney¹,³, Cindy Reichel⁹, Dog Aging Project Consortium‡, Noah Snyder-Mackler¹⁰, Joshua M. Akey⁵, Daniel E. L. Promislow¹¹, Frances L. Chen¹,³†, Elinor K. Karlsson¹,³,¹²*†
宠物犬与人类共享相似的生活环境,且衰老速度更快,是衰老研究中极具价值的转化模型。本研究利用“犬类衰老项目”(Dog Aging Project)中7627 只犬的基因组和表型数据(其中976 只犬完成了159 种血液代谢物和临床分析物的检测),构建了首个犬类全基因组关联研究(GWAS)目录。犬类与人类的血液性状定位到同源基因位点,表明相关通路具有高度保守性。品种血统可解释血液性状的显著变异,且对被毛类型等外观特征的选择具有多效性代谢效应。借助混种犬的嵌合血统特征和纵向死亡率数据,本研究识别出短寿品种中升高且可预测个体死亡风险的血液性状(包括球蛋白和钾),以及长寿品种中富集的保护性性状(如乙醇胺)。尽管部分与衰老相关的性状与生长激素通路有关,但多数性状独立于该通路,表明犬类衰老具有多因素特性。这些发现确立了犬类作为转化系统的地位,为识别与延长健康寿命相关的衰老遗传决定因素和生物标志物提供了重要依据。
衰老的特征包括生理和认知功能的渐进性衰退、广泛的代谢重塑,以及对慢性疾病的易感性增加,给个体、家庭和医疗系统带来沉重负担[1-5]。这些变化的速度和轨迹在个体间存在巨大差异,反映了遗传、环境和生活方式因素的复杂相互作用[6,7]。包括全基因组关联研究(GWAS)在内的大规模遗传研究已识别出数千个与极端长寿和年龄相关疾病相关的基因位点[8-12]。然而,将这些位点与功能衰退联系起来的生物学机制仍知之甚少,限制了其向有效治疗策略的转化。
与仅针对长寿的研究相比,血液性状的遗传研究为理解衰老提供了互补且更具机制性的方法。许多循环血液性状与衰老相关通路相关。血清化学图谱和全血细胞计数等临床分析物既受遗传变异影响[13-16],也会随年龄相关的生理变化而改变[17-20],并可作为疾病风险的生物标志物,反映与病理相关的过程[21-23]。血浆代谢组包含数千种反映代谢活动的小分子,可预测犬类和人类的死亡率、衰弱程度及功能衰退[24-28]。识别血液性状水平和纵向轨迹个体差异背后的遗传变异,有望为衰老相关疾病开发新的干预手段—— 例如低密度脂蛋白胆固醇(LDL-C)与他汀类药物治疗心血管疾病的研发[29]。
在人类研究中,识别影响衰老的可修饰代谢或环境因素受限于人类较长的寿命[30]。宠物犬(又称伴侣犬)具有许多与人类衰老相似的特征:其衰老轨迹受遗传、环境和生活方式因素影响而具有异质性[31-33],但衰老速度更快,寿命约为9-15 年[34-36]。作为家庭动物,犬类与主人经历相似的饮食、社会和环境暴露,且通常接受相当水平的医疗护理[37,38]。这种紧密的人犬关系使得收集高分辨率纵向数据成为可能,包括兽医临床记录、血浆代谢组学分析以及主人报告的生理和认知功能评估[39-43]。犬类与人类中,与衰老相关的指标(如血浆代谢物死亡率风险比)具有强相关性,证实了该系统的转化价值[44]。
与人类相似,犬类的血液性状随年龄、性别和体型而变化,但同时也受品种影响[43,45-47]。现代犬品种是过去175 年内通过严重的种群瓶颈和针对物理及生理性状的持续选择性育种形成的近交种群[48-51]。在美国,大多数犬为纯种犬(single-breed)或具有多个品种血统的混种犬(mixed-breed)[52]。这种特殊的种群历史改变了等位基因频率谱,使得在规模小于人类研究的队列中也能开展高效力的GWAS [53,54]。
纯种犬的寿命通常短于混种犬[34],部分大体型品种的寿命尤其短暂。品种间成年体型的变异与生长激素/ 胰岛素样生长因子1(GH/IGF-1)通路中的基因密切相关[55-57]。在啮齿动物模型中,该通路的调控会对衰老产生深远影响[58-61],这引发了一种推测:对GH/IGF-1 变异的选择可能导致部分犬品种寿命缩短[62]。然而,仅体型无法完全解释品种间的寿命变异[63,64]。美国大多数混种犬具有四个或更多品种的血统,体型跨度广泛[52],这形成了一个自然实验,可将体型的影响与其他品种相关遗传因素对衰老的作用区分开。
本研究利用“犬类衰老项目”(Dog Aging Project)中犬类独特的品种血统、环境多样性和深度纵向表型数据,识别与寿命变异相关的血液性状,并评估其与人类衰老的相关性。
犬类衰老项目
“犬类衰老项目”是规模最大的研究犬类衰老相关生物学、遗传和环境因素的项目[65]。截至2025年12月,该项目已招募52705只犬。本研究分析了2023年底前招募的47444只犬中具有遗传数据的7627只犬的子集。主人通过唾液拭子收集DNA样本,进行测序(平均测序深度0.79x±0.41x;范围0.14x至5.36x),并与CanFam4.0参考基因组比对[66]。基因型的估算来自包含1929只深度测序(12.95x至31.06x)犬类和相关犬科动物[67]。最终数据集包含29089701个双等位基因单核苷酸多态性(SNP),其中9871985个为常见SNP(次要等位基因频率≥0.01)。此前研究已表明,低覆盖度测序结合填充技术可达到与芯片相当的基因型准确性,同时产生10-50倍更高的标记密度,足以捕获大多数常见变异[52]。
遗传决定的品种血统
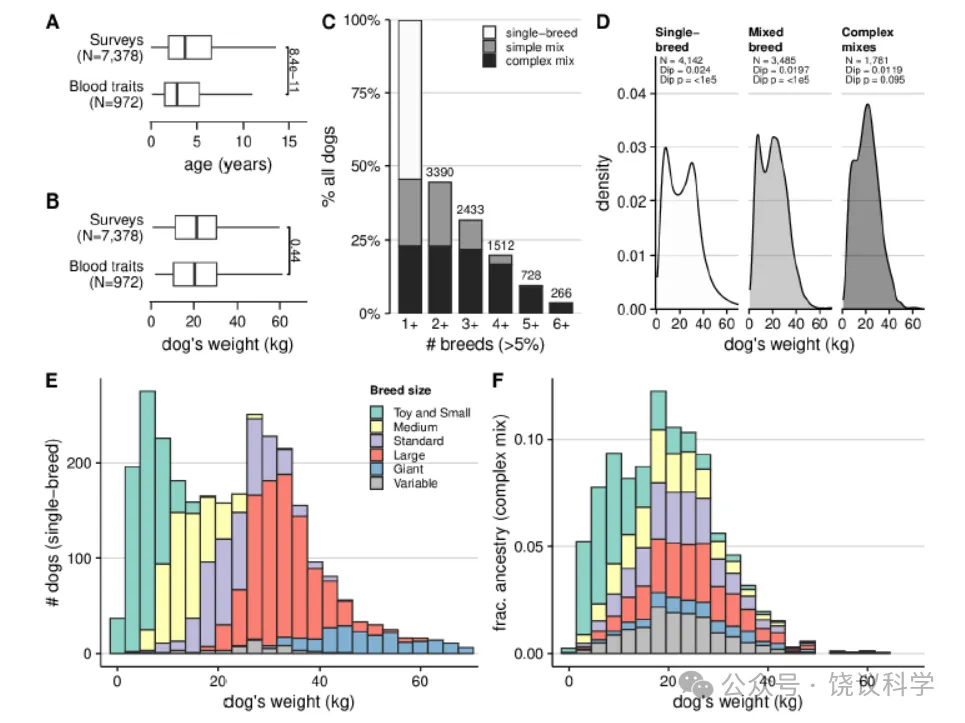
测序犬的入组年龄从数周至21岁不等(均值±标准差,4.6±3.5岁),比整个入组队列更年轻(6.8±4.3岁)(图1A、B;图S1A)。约一半为雌性(48.8%),82%已绝育,与美国宠物犬种群特征相似(图S1B)[68]。我们通过监督混合分析(supervised admixture analysis),以包含全球公认的360个品种中筛选出的109个常见品种、乡村犬和狼组成的参照组为依据,估算品种血统[69]。根据主人报告的信息将犬分为纯种或混种——此前研究已证实该分类方式可靠[52,69],且可将参照组中未包含的品种纳入数据集。
图1 研究种群的人口统计学和遗传结构
(A)具有血液性状数据的犬比整个入组种群更年轻;(B)体重无差异。(C)近一半的犬(45%)具有可检测到的多个品种血统(>5%),包括简单混种(任一品种血统占比> 40% 的犬)和更复杂的混种。(D)按血统复杂性分层的体重分布显示,纯种犬和所有混种犬的体重呈双峰分布,而复杂混种犬不呈双峰分布(采用哈蒂根双峰检验验证单峰性,100,000 次bootstrap 重复)。(E)在纯种犬中,体重与品种标准体型相关。(F)在复杂混种犬中,体型与品种血统的关系不太明确—— 大型品种血统可能出现在小型犬中,反之亦然,且极少出现体型极小型或极大型的犬。
混种犬的品种血统比例与纯种犬大致相似。总体而言,45.7% 的测序犬为混种犬,其中一半(51%)为“复杂混种”(任一品种的血统占比不超过40%)(图1C)。纯种犬中常见的品种在混种犬的血统中占比也更高(斯皮尔曼相关系数ρ=0.70)。在最常见的品种中,金毛寻回犬(golden retrievers)和拉布拉多寻回犬(labrador retrievers)的血统在纯种犬中更普遍,而美国比特斗牛梗(American pit pull terriers,)、贵宾犬(poodles)、玩具贵宾犬(toy poodles)和吉娃娃(Chihuahuas)的血统在混种犬中更常见(图S2)。
品种相关的种群结构影响成年体重分布,导致其偏离随机交配种群中预期的近似正态分布[70]。在纯种犬中,极小型和极大型品种的受欢迎程度使得体重分布呈双峰分布(n=4163;哈蒂根双峰检验,p0.2)[83,84]。遗传力越高的性状,越可能存在至少一个全基因组显著性位点(逻辑回归OR=9.5,p=0.0036),且总体显著性位点数量更多(拟泊松回归β=3.1,p=2.6×10⁻⁸)(图2B)。体重的结果清晰体现了这一模式,共产生33个不同的关联(图2C),包括已报道的IGF1(p=9.0×10⁻⁵⁰)、HMGA2(p=2.35×10⁻⁴⁹)和LCORL(1.33×10⁻⁴⁵)基因关联[52,55,56,77,85]。我们还识别出一个此前未报道的与microRNA-497邻近的关联(p=1.3×10⁻¹⁴),该microRNA是胰岛素信号基因邻近的IGF1R调控因子[86,87]。
其余153 个调查问题产生76 个显著性关联,其中67% 无显著性关联位点,22% 有1 个显著性关联位点,11% 有2 个及以上。除体重外,调查性状的关联显著性通常低于血液性状(图2C)。最强关联来自调查问题“与熟悉的犬玩耍玩具时表现出攻击性”(GWAS p=4.1×10⁻¹²),该位点在人类中与循环性激素结合球蛋白水平相关(rs79391862-A,p=2×10⁻¹²¹)[88]—— 这一表型主要反映代谢和内分泌状态,但可能间接影响行为。
相比之下,血液性状比调查性状产生更多、更显著的关联(图2C)。在159种血液性状中,识别出189个全基因组显著性关联(平均每种性状1.19±2.35个),其中82种血液性状(52%)至少有一个关联。最强关联来自血浆代谢物,且定位到具有明确生化关联的基因。例如,我们验证了此前报道的GPT基因与循环丙氨酸转氨酶水平的关联[54]。最显著的关联是代谢物3 -羟基异戊酸与MCCC2基因中的SNP——MCCC2 基因编码一种防止该代谢物积累的酶。
血液性状的位点异质性和多效性
犬类血液性状的遗传结构显示出广泛的位点异质性和多效性,与人类代谢组学和血液学GWAS中观察到的模式一致[29,89,90]。在82种至少有一个显著性关联的性状中,37种映射到多个独立位点,其中代谢物3 -羟基异戊酸(17个位点)、5 -甲基胞苷(14个位点)和S -甲基半胱氨酸(13个位点)的位点数量最多。189个全基因组显著性位点中,13 个与多种血液性状相关。
多效性最强的位点是13号染色体上包含ALB(编码白蛋白)的95kb区域。该位点的变异与色氨酸水平(p=1.8×10⁻⁶⁰)、白蛋白/球蛋白比值(p=1.1×10⁻¹⁷)、白蛋白水平(p=1.9×10⁻¹³)和球蛋白水平(p=2.7×10⁻⁸)相关。这一模式具有生物学合理性:约90%的循环色氨酸与白蛋白结合[91,92],因此改变白蛋白浓度或结合能力的遗传变异会同时影响色氨酸的可利用性。与这一机制一致,ALB位点还与已知结合白蛋白的代谢物相关(图S6),包括L -犬尿氨酸(p=5.6×10⁻⁴⁹)、吲哚- 3 -乙酸(p=4.1×10⁻²⁹)、吲哚- 3 -乳酸(p=4.2×10⁻²⁷)和吲哚- 3 -丙酸(p=2.3×10⁻¹³)[93,94]。
犬类与人类血液性状位点的重叠
犬类血液性状最显著相关的位点与人类研究中涉及的位点同源,表明存在共享的生物学机制,突显了犬类作为转化模型的价值。我们将犬类关联与25 项研究中同源人类GWAS 位点(±250kb)的报道进行了比较[95-120](数据S4)。在所有181 个犬类位点中,27% 与人类中相同或直接相关性状的关联重叠。当仅关注犬类最显著的信号时,这一重叠率大幅上升:在犬类中p0.05);标注10 个最强效应。(E)20 种受品种影响最大的性状在年龄、品种、性别和体重之间的变异分配(堆叠条形图);均无显著的体重效应。(F)临床分析物的犬类与人类效应比较。左侧:已发表的人类风险比;右侧:犬类中品种对变异的解释度。
体重仅能解释所有血液性状变异的一小部分,部分原因是品种与体型的强相关性(图3A)[71]。多重共线性诊断显示,这种相关性通常不会掩盖模型中体重的独立效应(方差膨胀因子VIF=2.72)。然而,对于部分血液性状,纳入品种后会显著减弱体重的表观效应。例如,在不含品种的模型中,体重解释14.5% 的肌酐水平变异(肌酐是反映肌肉总量的代谢物)[124];而纳入品种后,体重的解释度几乎为零(0.4%),品种的解释度为38.3%。在不含品种的模型中,体重对14 种血液性状的变异解释度超过5%,纳入品种后,体重效应平均降低94.1%±6.7%,剩余解释度仅为0.01%-1.4%(图3C)。
总体而言,96 种血液性状的变异可由品种显著解释(FDR p1.9)和钾升高(HR>1.4)均会增加死亡风险[143,144]。
8 种血浆代谢物(4 种风险相关,4 种保护性)在联合分析中被证实相关(图5E、图S12)。风险相关代谢物包括:辛二酸suberic acid(一种指示线粒体应激的二元羧酸,随人类年龄增长而积累[145])、二甲基甘氨酸dimethylglycine,(肾功能障碍的标志物[146],与全因死亡率增加相关[147,148]),以及乙基丙二酸ethylmalonic acid和琥珀酸succinate(均为线粒体功能障碍的标志物)。保护性代谢物包括:乙醇胺ethanolamine,(通过增强自噬延长酵母、果蝇和哺乳动物细胞系的寿命[149])、脱氧肉碱deoxycarnitine(肉碱通路通量的生物标志物,其水平降低与衰弱、胰岛素抵抗、心血管疾病和肌肉无力相关[150,151]),以及胍乙酸guanidinoacetate(肌酸的直接前体,肌酸水平降低与肌肉减少症、疲劳和认知衰退相关[152-154])。
最强的保护性候选物之一是羟脯氨酸hydroxyproline —— 它并非人类中典型的衰老生物标志物。羟脯氨酸主要源于胶原蛋白降解,可能反映富含胶原蛋白组织的周转[155]。长寿品种血统占比更高的犬,羟脯氨酸水平更高,且个体犬中羟脯氨酸水平与较低的死亡风险相关。