人类寿命存在极限的证据自然
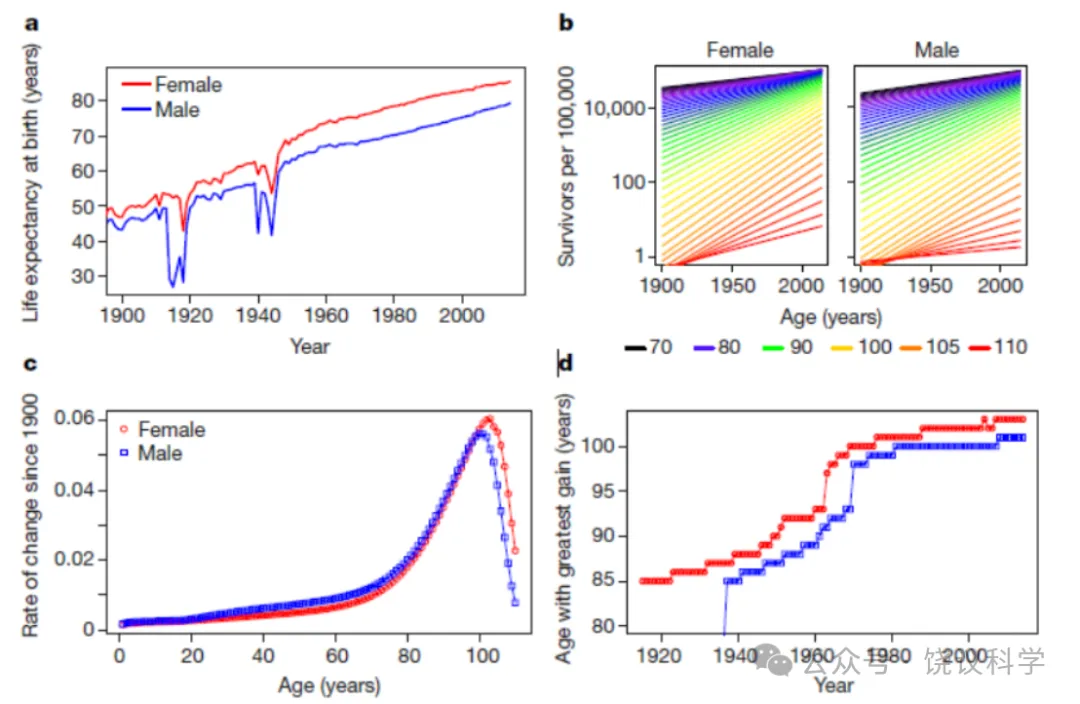
在技术进步的推动下,自十九世纪以来,人类预期寿命已大幅增长。人口学证据显示,老年死亡率持续下降,最高死亡年龄不断上升,这可能逐步延长人类寿命。加之观察到各种动物物种的寿命具有可塑性,并可通过遗传或药物干预延长,这些结果引发了以下观点:长寿可能不受严格的、物种特异的遗传限制。在此,通过分析全球人口统计数据,我们表明,百岁之后,随年龄增长的存活率改善趋于下降,且自20世纪90年代以来,全球最年长者的死亡年龄未曾增加。我们的结果强烈表明,人类的最大寿命是固定的,并受自然约束的限制。
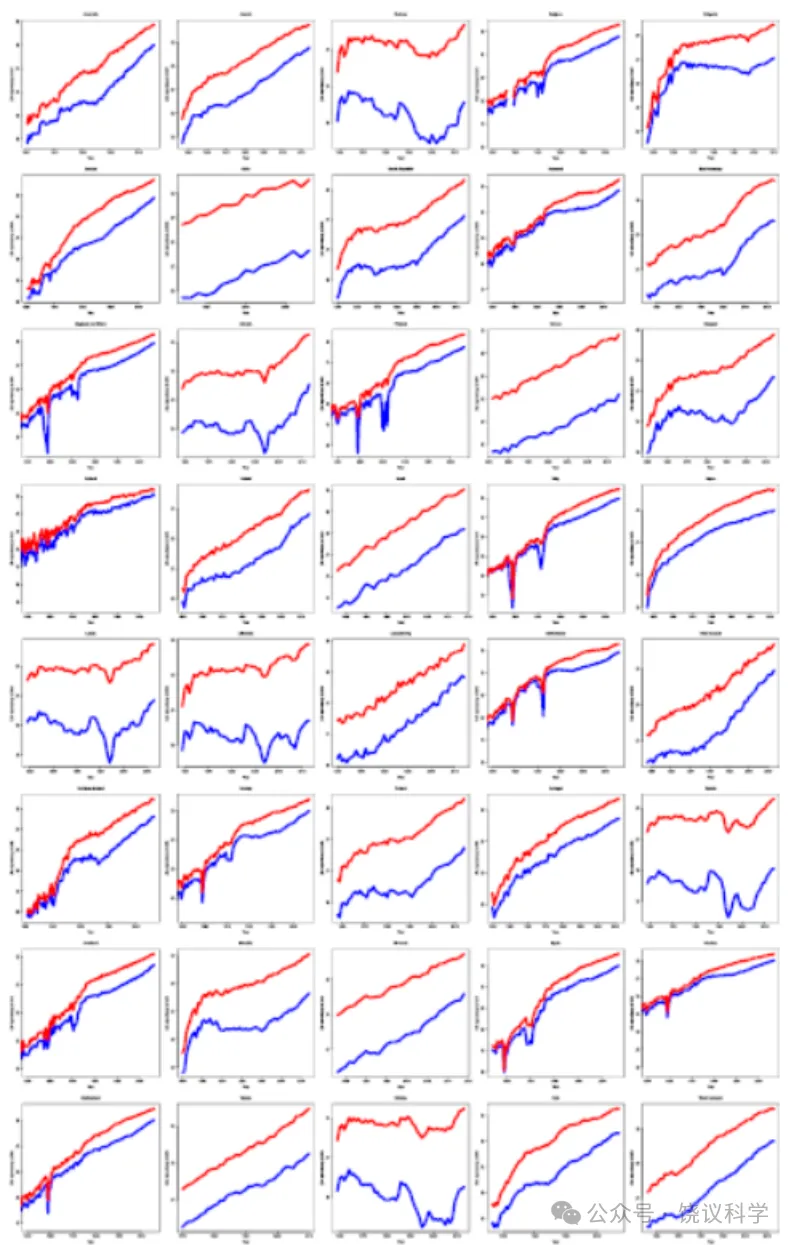
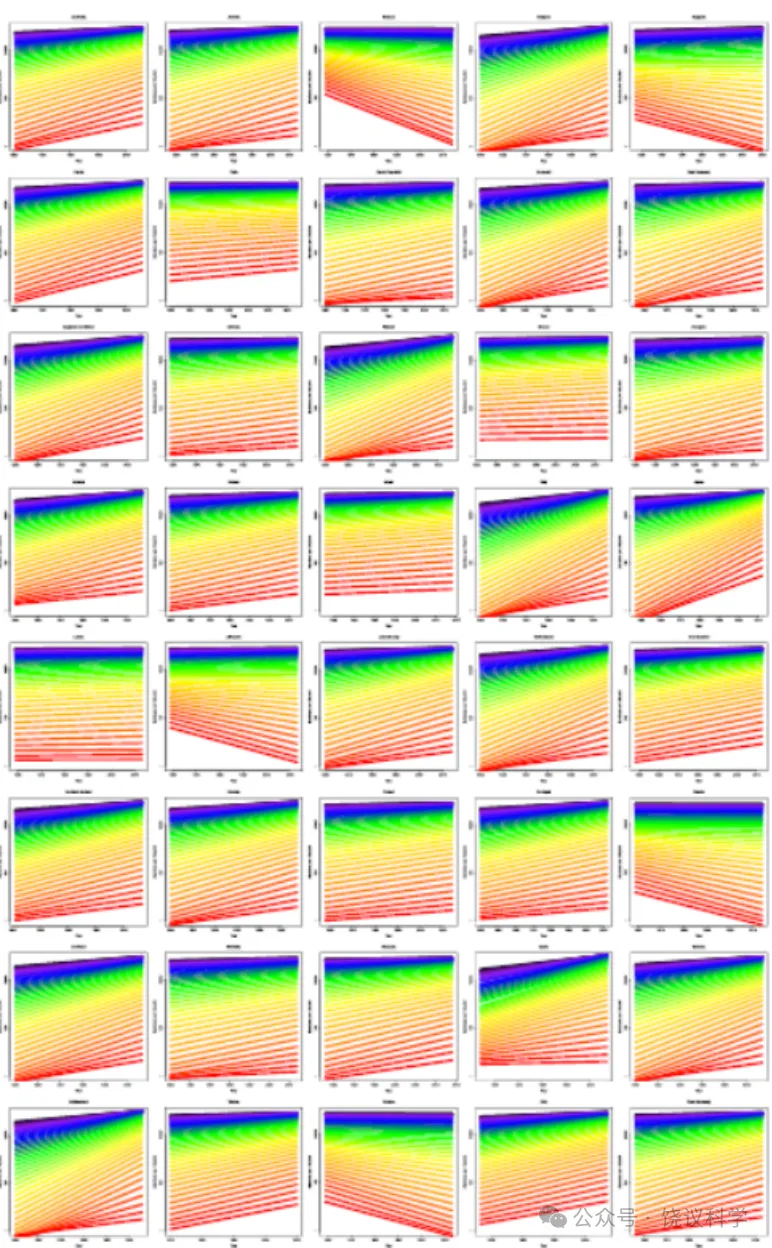
与平均寿命相反,最大寿命通常被认为是一个物种的稳定特征。对于人类,报告的最高死亡年龄通常设定为122岁,即珍妮·卡尔芒(Jeanne Calment)的死亡年龄,她仍然是有记载以来最长寿的人类个体。然而,一些证据表明最大寿命并非固定不变。在模式生物中的研究表明,最大寿命具有可塑性,可受遗传和药理干预影响。在瑞典,基于一系列关于人类寿命上限的长期可靠信息,发现报告的最高死亡年龄从19世纪60年代的约101岁上升至20世纪90年代的约108岁。据此,作者认为,这一发现驳斥了人类寿命固定不变且不随时间推移而改变的普遍论断。事实上,支持人类最大寿命非固定不变的最有力论据是,上个世纪大多数国家预期寿命的持续增长。图1a展示了法国(一个拥有高质量死亡率数据的国家)的这一增长,但大多数其他发达国家也发现了非常相似的模式(扩展数据图1)。因此,人们考虑了一种可能性,即死亡率可能进一步下降,打破任何预设的人类寿命界限。
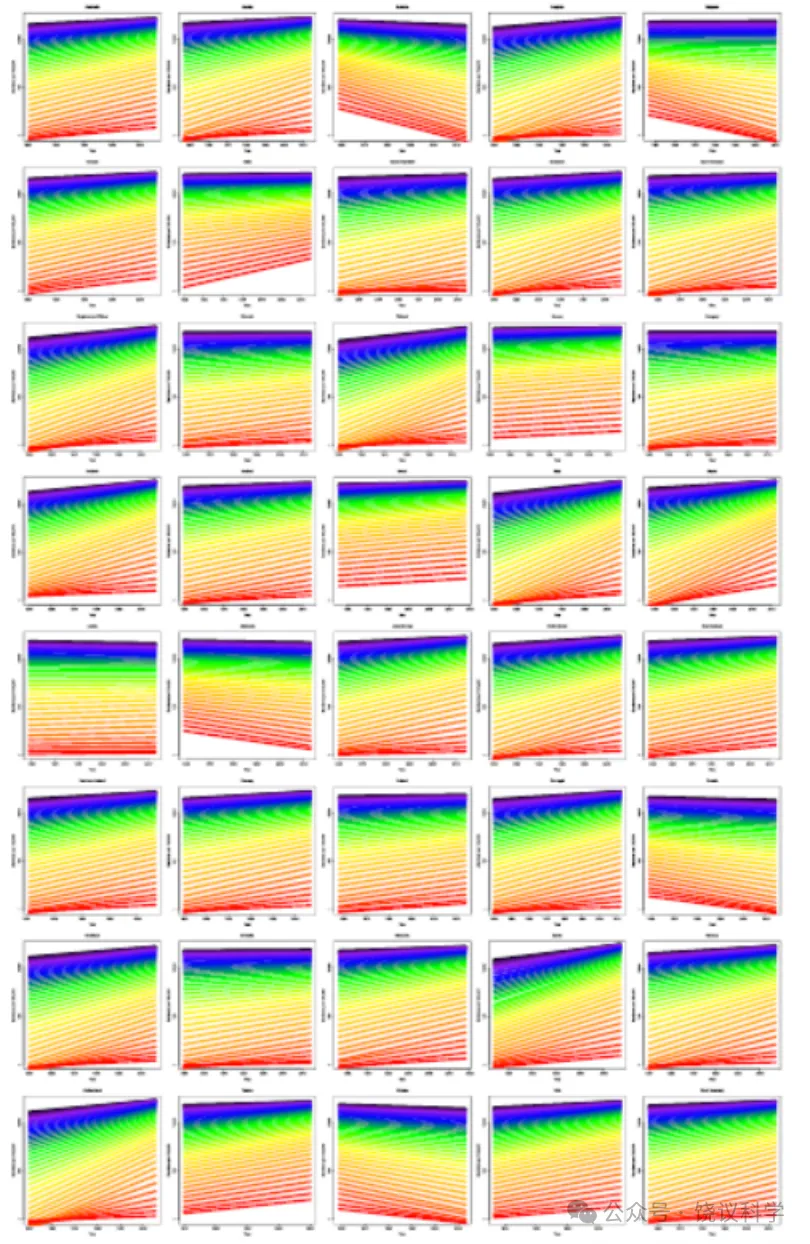
正如人类死亡率数据库的数据所示,历史上预期寿命的许多增长可归因于早期寿命死亡率的降低。然而,更近期的数据显示出老年死亡率下降的证据,每个出生队列存活至老年的比例随日历年份增加。在法国,每10万人中存活至老年(70岁及以上)的人数自1900年以来有所增加(图1b),这表明人类预期寿命持续增长。这一模式在数据库所包含的其他40个国家和地区中非常相似(扩展数据图2、3)。然而,存活率改善的速率在极高年龄水平处达到峰值后下降(图1c),这表明老年死亡率降低的收益递减,并提示人类寿命可能存在极限。在其他发达、低死亡率国家也发现了相同的模式(扩展数据图4)。然而,我们考虑了一种可能性,即存活率增长最快的年龄随日历年份推移而增加;也就是说,存活至老年率增长速率的峰值将随时间向右移动。为验证这一点,我们绘制了该峰值出现的年龄与日历年份的关系图(图1d)。结果表明,存活率改善最大的年龄似乎在1980年左右趋于平稳。在数据库中41个国家中的88%中观察到了类似模式(扩展数据图5)。总之,这些发现表明(但未证明)人类寿命可能存在自然极限。为进一步研究这一观点,我们将注意力从老年死亡率转向人类最大寿命本身,并考察了全球最年长个体的死亡年龄。
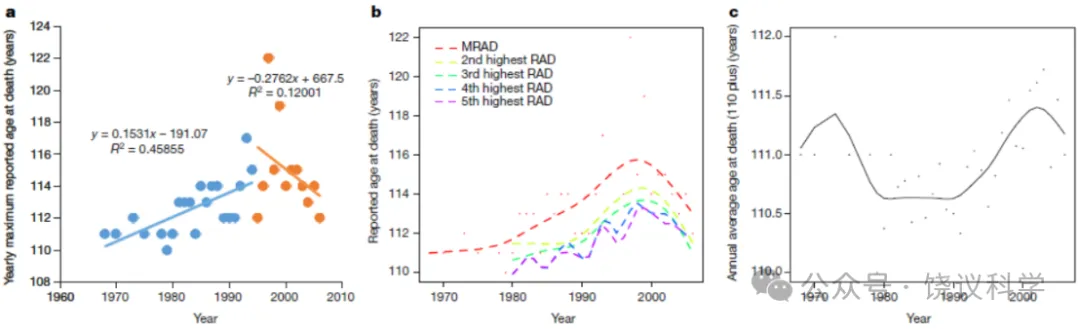
我们首先绘制了法国、日本、英国和美国(这些国家在国际长寿数据库(International Database on Longevity,IDL;n = 534,1968–2006年)中记录了最多的超级百岁老人(年龄≥110岁))的每年最高报告死亡年龄(maximum reported age at death,MRAD)。如图2a所示,尽管死亡年龄在20世纪70年代至90年代初间迅速上升,但在1995年左右达到平台期,接近1997年——即珍妮·卡尔芒去世的年份。我们将数据分为两组(1968–1994年和1995–2006年),并使用线性回归对每组进行建模。结果表明两组之间存在趋势断点。1995年之前,MRAD每年增加0.15岁(r = 0.68,P = 0.0007);然而,1995年之后,MRAD不再显著增加,实际上每年略有下降0.28岁(r = -0.35,P = 0.27)。当我们考虑来自另一个独立资源——老年学研究组织(Gerontological Research Group,GRG;http://www.grg.org/)的MRAD记录时,我们观察到类似的趋势——在1972–1994年间每年增加0.12岁(r = 0.71,P = 0.0002),随后在1995–2015年间每年略有下降0.14岁(r = -0.36,P = 0.70)(扩展数据图6)。这些结果表明,尽管MRAD在20世纪90年代之前一直在增加,但此后未再观察到进一步增加;人类年度MRAD已稳定在114.9岁(95%置信区间:113.1–116.7岁)。为逼近人类寿命的绝对极限,我们将MRAD建模为泊松分布;我们发现,在任何给定年份中MRAD超过125岁的概率小于万分之一。
我们结果的一个潜在混杂因素是报告的MRAD病例数量相当少,这可能将这些结果仅仅解释为波动。为提供一个稳健的统计模型以加强观察到的模式,我们考虑了几组高报告死亡年龄(HRAD),即最高RAD(MRAD)以及第二至第五高的RAD(图2b;数据汇总自IDL)。所有序列均表现出与MRAD相同的模式。值得注意的是,甚至这些超级百岁老人的年度平均死亡年龄自1968年以来也未增加(图2c)。
因此,与先前关于人类寿命可以进一步延长的建议相反,我们的数据强烈表明寿命是有限的。过去,其他人也曾提出人类寿命是有限的。例如,1980年,Fries认为,由于寿命有限,增加过早死亡的预防将导致发病率的压缩。然而,他关于寿命有限性的论点——即百岁老人或最高报告死亡年龄缺乏可检测的增长——在当时虽然正确,但后来已被推翻。十年后,Olshansky等人基于死亡率的假设性降低估计了人类寿命的上限,得出结论认为出生时预期寿命不会超过85岁。与Fries一样,Olshansky等人也基于经核实的最长寿个体年龄缺乏增长而提出了生命的生物学极限。然而,正如他们所指出的,数据不足使他们无法得出明确结论。如今,二十多年过去了,这类数据正变得可用。尽管本研究中超级百岁老人死亡年龄的数据仍存在噪声且样本量较小,但我们认为图2中观察到的轨迹令人信服,且我们的结果强烈表明人类寿命存在自然极限。
这种人类寿命极限的生物学原因可能是什么?关于衰老是自然选择直接作用下进化出的有目的、程序化事件序列以导致死亡的观点,现已基本被否定。相反,看似自然的极限是早期生命事件(如发育、生长和繁殖)的固定遗传程序的无意副产物。寿命的极限很可能由基因组中编码的一套物种特异的、长寿保障系统所决定,这些系统对抗这些无意副产物——其中可能包括将遗传信息转化为细胞功能时的固有缺陷。要将人类寿命进一步延长至超越这些长寿保障系统所设定的极限,则需要超越改善健康寿命的干预措施,其中一些目前正在研究中。尽管没有科学理由表明这些努力不可能成功,但可能性本质上受到共同决定物种特异性寿命的大量遗传变异的约束。
图1预期寿命和老年生存趋势。 a,每一年份人口的出生时预期寿命。法国预期寿命在20世纪和21世纪初有所增加。b,存活至老年的人口比例的回归分析表明,自1900年以来存活率有所增加,但100岁及以上年龄的增长率似乎较慢。c,绘制变化速率(对数转换数据回归所得系数)表明,生存率增长在100岁左右达到峰值,之后迅速下降。d,日历年份与过去100年中生存率增长最快的年龄之间的关系。生存率增长最快的年龄在过去一个世纪中有所增加,但其上升速度一直在减慢,似乎已趋于平稳。
图2 超级百岁老人的报告死亡年龄。 所有数据均收集自IDL数据库(法国、日本、英国和美国,1968–2006年)。a,每年最高报告死亡年龄(MRAD)。线条表示线性回归函数。b,每年第一至第五高报告死亡年龄(RAD)。虚线是使用三次样条对RAD的估计。c,每年最高报告死亡年龄(MRAD)、第二高至第五高RAD以及该组(MRAD及第二至第五高)平均值的年度变化。所有序列在20世纪90年代中期左右趋于平稳。
扩展数据图1 | 自1900年(或数据可用的最早年份)以来40个国家和地区的预期寿命随时间变化。 总体上呈正趋势;日本预期寿命似乎趋于平稳,但许多其他国家的增长似乎未见减弱。数据代表每个地区的总人口,苏格兰除外(仅代表平民人口)。颜色方案与图1a相同。
扩展数据图2 | 40个国家和地区女性存活至老年的人口比例。 数据代表每个地区的总人口,苏格兰除外(仅代表平民人口)。颜色方案与图1b相同。
扩展数据图3 | 40个国家和地区男性存活至老年的人口比例。 数据代表每个地区的总人口,苏格兰除外(仅代表平民人口)。颜色方案与图1b相同。