研究解析减数分裂异染色质区重组抑制的分子机制中国科学报
在国家自然科学基金等项目资助下,华南农业大学生命科学学院教授王应祥团队系统解析了ATXR5/6蛋白抑制减数分裂异染色质区重组形成的分子机制。相关成果近日发表于《植物学报(英文版)》(JIPB)。
减数分裂重组对世代繁衍、物种多样性维持及作物遗传育种具有重要意义。交叉(crossover,CO)作为同源染色体重组交换遗传信息的产物,在染色体上呈非随机分布。约80%以上的重组事件集中在约25%的基因组区域(即重组热点区域),且倾向于发生在常染色质区,而在近着丝粒的异染色质区则受到抑制。然而,这一分布规律背后的调控机制尚不清晰。
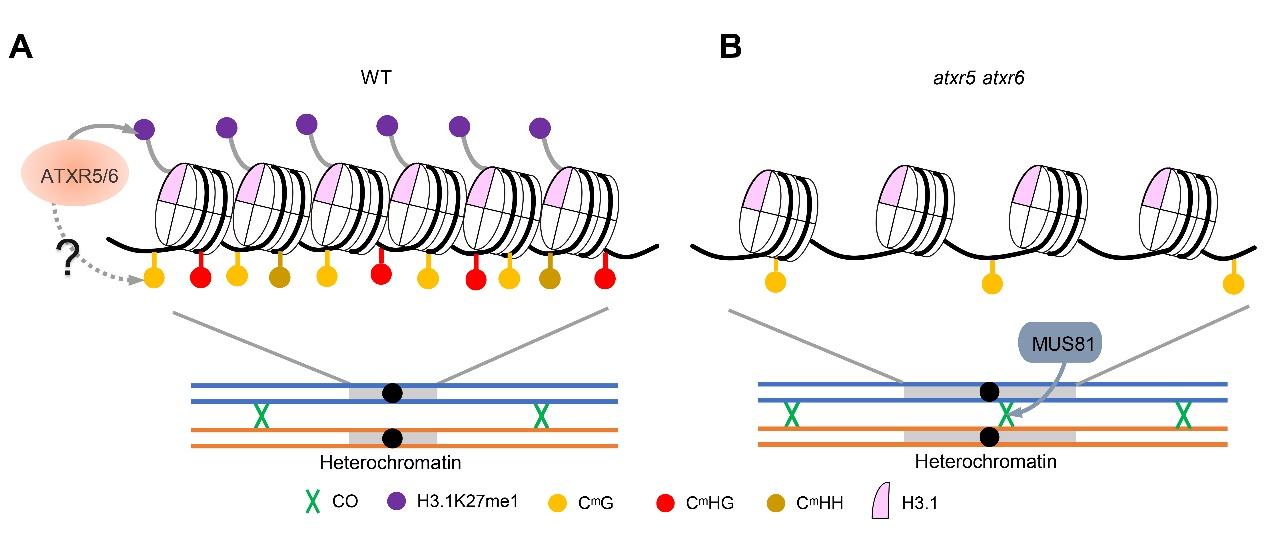
ATXR5/6在调控减数分裂异染色质重组中的作用模型。研究团队供图
已有研究表明,H3K9me2、DNA甲基化、组蛋白变体等表观修饰参与重组调控。植物特有的H3K27me1主要富集于异染色质,由植物特异的ATXR5/6(ARABIDOPSIS TRITHORAX-RELATED PROTEIN 5/6)催化。此前研究仅知ATXR5/6在体细胞中发挥维持异染色质压缩、沉默转座子的功能,但其是否参与减数分裂重组的调控尚不明确。
该研究发现,ATXR5和ATXR6通过调控异染色质凝缩及其修饰来抑制减数分裂异染色质中CO的形成。在atxr5 atxr6双突变体中,减数分裂CO数目显著增加,回补实验可完全恢复表型,证实ATXR5/6对减数分裂CO形成起负调控作用。通过遗传分析与重组标记蛋白的免疫荧光实验,研究证明ATXR5/6突变可能不影响DNA双链断裂的形成,但能够同时促进干涉敏感型和干涉不敏感型两类重组通路。
进一步研究发现,在双突变体中,减数分裂异染色质区出现去凝缩现象,H3K27me1信号减弱,DNA甲基化特异性降低,同时近着丝粒区异染色质区的重组频率显著升高。与既有研究一致,ATXR5/6在体细胞中主要调控异染色质凝缩和基因/转座子沉默而不影响DNA甲基化。意外的是,在减数分裂细胞中,ATXR5/6对基因表达的影响很小,也不调控减数分裂功能基因的表达,却特异性维持异染色质的CHG甲基化与H3K27me1修饰。这一结果提示,ATXR5/6主要通过维持异染色质的表观修饰和结构来抑制重组,而非通过转录调控路径发挥作用。
此外,在atxr5 atxr6背景下进一步敲除重组因子MMS AND UV SENSITIVE 81(MUS81),会引发严重的营养生长和生殖发育缺陷。三突变体的减数分裂过程中出现着丝粒异常粘连和碎片化,导致多分体形成,花粉活力和结实率显著下降,表明着丝粒周边发生的异常重组无法得到正常修复。这揭示MUS81对于解除ATXR5/6缺失后异染色质区产生的异常重组中间体至关重要,有助于维持基因组稳态。
论文共同通讯作者、华南农业大学生命科学学院教授王聪指出,该研究拓宽了ATXR5/6在生殖细胞中区别于体细胞的表观遗传功能认知,并为突破作物异染色质区域的重组瓶颈、打破杂交育种中的连锁累赘、实现优异等位变异快速组合提供了重要的理论基础。