小脑参与语言?机器翻译
尽管已有大量证据表明小脑参与认知功能(包括语言加工),但其确切作用仍存在争议。本研究借助精准功能磁共振成像(precision fMRI)技术,系统刻画了小脑中对语言产生响应的脑区。我们在右后侧小脑鉴定出4 个跨感觉通路均对语言产生稳定响应的脑区。其中一个脑区横跨小脑 Crus I/II 及小叶 VIIb,对语言具有高度选择性,显著区别于各类非语言任务;其余三个脑区则表现为混合选择性,即同时对语言与非语言输入产生响应。与新皮层语言系统类似,该语言选择性脑区参与语言理解与产生过程,对语言加工难度敏感,且对社会性与非社会性句子均产生响应。最终,全部 4 个小脑脑区(尤以 Crus I/II/VIIb 区域最为显著)均与新皮层语言网络存在功能连接。我们提出,这些小脑区域构成扩展语言网络的组成部分:其中一个区域与新皮层语言网络高度同源,其余区域则可能负责整合来自多个新皮层网络的信息。
人类小脑虽仅占脑体积的 10%,却包含了大脑中近 80% 的神经元,其表面积接近大脑皮层的 80%,是计算能力极强的神经结构。传统观点将小脑与运动控制关联,而数十年的神经成像与神经心理学研究现已证实,小脑同样参与人类认知功能(包括语言),甚至可能在演化上促成了认知能力的出现。
小脑由一系列功能独立的亚区镶嵌组成,其中包括参与语言加工的区域(如小脑 Crus I/Crus II)。神经成像研究多次报道,右后侧小脑在多种语言任务中激活,包括动词生成、句子补全与被动理解等。类似地,该区域损伤可引发多样的语言缺陷,包括语法障碍、韵律异常、命名不能、失读、言语流畅度下降及元语言缺陷。然而,将这些零散发现整合成一套连贯的小脑语言功能理论,始终面临挑战。
小脑语言研究的进展受限于若干共性问题,部分问题在语言神经科学领域普遍存在,部分则为小脑研究所特有。首先,与早期新皮层语言脑区研究相似,多数用于研究小脑语言功能的任务未能将语言加工与知觉、运动及非语言认知过程分离。例如,动词生成或言语流畅度任务会混淆语言加工与一般性任务需求,且多数语言范式缺乏知觉或运动控制条件,导致难以对观测到的响应做出无歧义解读。其次,过往多数小脑成像与损伤研究依赖解剖定位与被试间平均分析,精准功能磁共振成像技术应用有限,难以判断不同研究观测到的是否为同一脑区。例如,多项研究报道右后侧小脑在语言加工及其他认知过程(如心理理论、执行控制)中激活,但缺乏被试内比较,无法确定小脑语言脑区是否与这些脑区重叠并支持非语言功能。
尤其在小脑语言研究中,研究通常仅考察一两种范式下的响应,导致其底层计算机制不明确。部分研究针对小脑在语言中的特定作用提出假说(如语言预测),并基于小脑环路模型展开验证,但未检测相关脑区对其他语言层面或非语言任务的响应。因此,这些研究无法区分目标脑区是执行语言预测还是更广泛的语言加工,也无法判断其计算是语言特异性还是领域一般性。另一些研究试图通过多领域任务组合或大量自然认知数据(如静息态数据)解析小脑整体组织,这类研究能区分不同小脑区域,但对特定领域内的功能解析较为粗糙。
最后,新皮层与小脑语言脑区的研究长期相互分离,绝大多数研究聚焦新皮层,导致小脑语言系统相对于新皮层环路的作用仍不明确。语言神经科学被割裂为皮层与小脑两大分支,阻碍了整合性理论的构建,而这对于全面阐释大脑语言加工机制至关重要。
本研究采用已成功应用于新皮层语言系统解析的研究范式,揭示小脑在语言加工中的作用。具体而言,依托大规模功能磁共振成像数据集(846 名独立被试、1033 次扫描、26 项实验),我们:1)采用成熟的语言功能定位范式,在小脑中搜寻语言响应脑区;2)刻画这些脑区相对于其他功能的语言选择性;3)针对语言选择性脑区,解析其语言加工的具体机制;4)比较小脑语言脑区与新皮层语言网络的相似性及功能连接。
右后侧小脑存在四个跨感觉通路的语言响应脑区
我们首先在大样本被试中探究小脑语言响应的拓扑分布。所有被试完成语言定位范式:阅读句子与知觉特征相似但无意义、无结构的非词串。语言 > 控制条件(句子 > 非词)靶向支持词汇识别、句法构建、语义组合的脑区,且可跨感觉通路、任务与语言泛化。
为在小脑中搜寻语言响应脑区,我们采用组约束被试特异性(GSS)分析方法。该方法与传统组分析类似,可搜寻被试间空间一致的激活区域,但关键在于能保留被试间功能脑区精确位置的个体差异 —— 这一点在小脑中尤为重要,因为小脑语言响应的拓扑分布在被试间差异显著。尽管存在个体差异,小脑语言响应的位置仍具备足够一致性,全脑 GSS 分析成功鉴定出4 个语言脑区,且全部位于右侧小脑。这一右半球偏侧化现象在全小脑语言响应体素中同样显著。
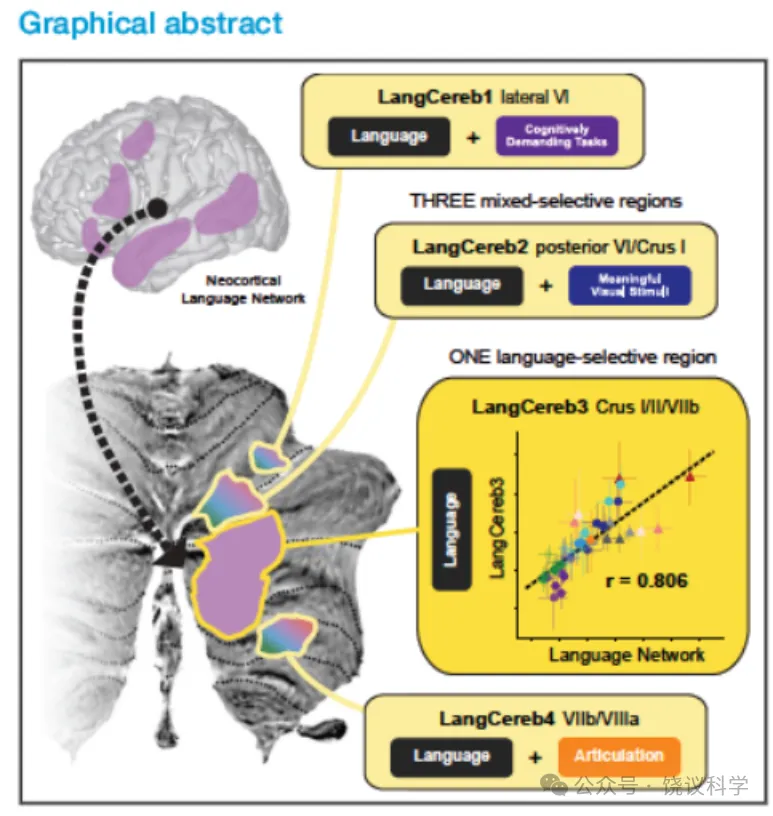
4个语言脑区均位于右后侧小脑,按从上至下顺序依次为:1)最小脑区,位于外侧小叶 VI(LangCereb1);2)横跨后侧小叶 VI 与 Crus I(LangCereb2);3)最大脑区,横跨 Crus I、Crus II 及小叶 VIIb(LangCereb3);4)横跨小叶 VIIb 与 VIIIa(LangCereb4)。这些脑区的位置与既往报道的小脑语言脑区部分重叠。我们在每名被试中定义功能感兴趣区(fROI),并采用独立数据检验其对语言的响应。结果显示,全部 4 个脑区对语言的响应均显著强于控制条件。
在新皮层中,同一语言网络同时支持阅读与听觉语言理解。因此我们进一步检验小脑语言响应是否同样跨感觉通路泛化。部分被试同时完成基于听觉的语言定位任务:聆听《爱丽丝梦游仙境》短文,对照条件为声学退化、无法理解的同一段语音。结果显示,阅读与听觉条件下,小脑激活的拓扑分布在被试内与组水平均高度相似。所有四个小脑语言脑区在阅读与听觉条件下均产生稳健响应,其中 LangCereb1(外侧小叶 VI)对视觉刺激偏好更强,LangCereb3(Crus I/II/VIIb)对听觉刺激偏好更强。上述结果表明,小脑语言脑区与新皮层同源脑区类似,参与抽象、跨感觉通路的语言输入加工。
一个小脑语言脑区(LangCereb3)表现出语言选择性,其余三个为混合选择性
我们进一步检验这些语言响应脑区是否具有语言选择性。对语言的选择性响应提示该脑区支持特异性语言计算,而同时对语言与部分非语言条件产生响应则提示其支持多重功能或执行共享计算。为此,我们检测小脑语言脑区对一系列非语言任务的响应。
我们将非语言任务分为五大类:1)言语发声:被试大声重复音节序列或手指敲击序列;2)高认知负荷执行功能任务:包含多种经典任务(如算术、空间工作记忆),均设置难易两种条件;3)音乐知觉:聆听器乐片段及缺乏层级结构的听觉控制刺激;4)动态视觉社会知觉:观看人脸、躯体视频及非生命物体视频;5)视觉事件语义:观看无声动画电影或静态事件图片,完成语义判断或知觉判断任务。
既往研究表明,新皮层语言脑区对非语言任务仅产生微弱响应,仅对有意义的非语言刺激产生中等强度响应,且显著弱于对语言的响应。在小脑中,仅 LangCereb3(Crus I/II/VIIb)表现出与之相似的选择性模式。该选择性表明此脑区不参与运动需求、一般性认知需求、层级结构加工、社会 / 交流信号或一般性意义刺激加工,与新皮层语言网络高度一致。
其余三个小脑语言脑区均对至少一种非语言任务产生与语言同等强度的响应:LangCereb1(外侧小叶 VI)对高认知负荷任务响应强烈,且对困难条件响应更强,兼具新皮层语言网络与多需求(MD)网络的功能特征,这一模式在新皮层中尚未见报道(新皮层中两大网络界限清晰);LangCereb2(后侧小叶 VI/Crus I)对有意义视觉刺激(事件图片、自然视频)响应强烈,支持跨语言与图片模态的语义加工,同时对高执行负荷任务产生响应;LangCereb4(VIIb/VIIIa)对运动任务响应强烈,言语发声条件下的响应数值上强于手部运动条件,同时类似 LangCereb1,对高认知负荷任务响应更强,呈现运动 / 发声控制与 MD 网络样的混合特征。
我们通过两项补充分析量化这些脑区的语言选择性及与邻近网络的重叠程度。首先,计算所有非语言任务下小脑与新皮层语言脑区的整体语言选择性:选择性定义为脑区对语言的响应与次高非语言条件响应的差值(经两者之和标准化)。结果显示,新皮层语言网络的语言选择性显著高于所有小脑语言脑区,对语言的响应强度是非语言任务的两倍;LangCereb3(Crus I/II/VIIb)是小脑中选择性最高的脑区,但仍低于新皮层网络;其余三个小脑脑区的选择性为 0(即至少一种非语言条件引发的响应等于或高于语言响应)。该结果提示这三个脑区并非仅执行语言计算,有待进一步实验解析其功能。
其次,评估小脑语言脑区与三个邻近认知与运动控制网络(心理理论 ToM 网络、MD 网络、言语发声网络)的可分离性。结果显示,LangCereb1–4 的语言活动峰值均可与 ToM、MD 及发声峰值分离。重要的是,这种可分离性并非必然存在:小叶 IX 即为反例,其语言定义体素(voxels)与 ToM 定义体素的功能谱完全一致,提示该区域最活跃的语言体素同时也是最活跃的 ToM 体素。
语言选择性小脑脑区(LangCereb3)加工句子水平语义,对语言加工难度敏感,且对社会性与非社会性语言均响应
确认 LangCereb3(横跨 Crus I/II/VIIb)具有语言选择性后,我们探究该脑区支持的具体语言计算。我们聚焦此语言选择性脑区,因其功能更可能与特异性语言过程相关;同时报告非选择性脑区的响应,但其功能解析留待未来研究。
一项广泛用于解析新皮层语言脑区功能的范式采用句子及多种语言退化条件,可分离语言加工的核心成分:词汇提取(识别单个词)、句法结构构建(确定词间结构关系)、语义组合(基于词汇与句法依赖构建句子水平表征)。该范式包含:有意义合语法句子(需全部三种成分)、无关联词汇表(仅需词汇提取)、合语法但无意义的荒诞句(仅需句法构建)、非词表(无需任何成分)。
所有新皮层语言脑区均对句子响应最强,对非词表响应最弱,对词汇表与荒诞句的响应居中,提示新皮层语言网络支持词汇提取与句法构建;且对句子的响应显著高于词汇表与荒诞句响应之和,提示其额外支持语义组合。与之不同,LangCereb3(Crus I/II/VIIb)对句子产生强烈响应,而对三种语言退化条件的响应均较低(词汇表与荒诞句的响应强度与非词表无显著差异),提示该脑区主要支持组合语义加工;事实上,LangCereb3 对组合语义加工的敏感性略高于新皮层语言网络。
我们进一步探究 LangCereb3 是否在语言产生中同样支持组合语义加工。被试需通过口语完成三种任务:描述图片事件产生句子、命名物体图片产生词汇表、朗读单音节非词表;另一组被试完成打字版任务。与理解结果一致,新皮层语言脑区对句子产生响应最强,词汇表产生居中,非词表产生最弱;而 LangCereb3(Crus I/II/VIIb)仅对句子产生强烈响应,对词汇表与非词表产生的响应均较低,口语与打字条件下结果一致,与其理解特征相符。
三个非选择性小脑语言脑区在理解与产生中呈现多样的功能特征,无任何一个完全匹配新皮层语言网络。
为更精细评估 LangCereb3(Crus I/II/VIIb)的响应如何受语言特征调节,我们检测其对 1000 个多样化句子的响应。首先,LangCereb3 与新皮层语言网络的响应谱高度相关,句子在新皮层网络引发强响应则在 LangCereb3 中同样引发强响应。事实上,两者间的响应相似性与各自脑区内的被试间相似性无显著差异。
随后,我们考察 LangCereb3 的响应如何受句子水平加工难度相关特征调节 —— 这类特征已知可调节新皮层网络响应。部分特征由语言模型自动提取,部分通过独立被试的行为评分获得。结果显示,LangCereb3 的响应受多数特征调节,包括由 GPT2-xl 计算的句子预测性(惊奇度),低预测性句子引发更强响应。更重要的是,控制惊奇度后,LangCereb3 与新皮层语言网络对加工难度相关特征的敏感性模式高度相似。三个非选择性小脑语言脑区的响应同样受这些特征调节,但响应模式与新皮层网络的相似性更低。上述结果表明,LangCereb3 不仅对语言刺激产生粗略响应,还对调节新皮层语言响应的特征表现出精细敏感性。