为什么我们都习惯用右手?DeepTech深科技
黑猩猩会用某一只手砸开坚果,但你很难说整个种群偏爱哪只手;而无论在东京、内罗毕,还是亚马孙雨林深处的村落,每十个人里大约有九个会用右手握笔、持刀、挥锤。你会用右手拧瓶盖、系鞋带、写字,这些习以为常的动作背后,却藏着一个人类演化中最顽固的谜题:为什么我们压倒性地偏向右手,而猿类就算会用工具,也没在群体层面形成同样的倾向?
数十年来,在这个问题上,遗传学、神经科学、发育生物学各自给出过线索:工具使用、投掷、语言、抱婴、社会竞争……几十种假说并存,却始终拼不出完整答案。
与其再提出一个新假说,不如把所有旧假说一次性放上同一架天平称一称。近日,一项牛津大学(University of Oxford)主导的新研究,终于把这个问题的答案,指向了人类演化中两件最具标志性的事件:直立行走,以及大脑的急剧扩张。
(来源:DOI: 10.1371/journal.pbio.3003771)
当用手偏好的强度和方向被拉到极值
人类的右手偏好是一种近乎普适、且高度稳定的现象。在已被记录的人类文化中,约九成的人惯用右手;基于考古证据,有学者认为,这一比例至少自新石器时代以来就保持稳定,也有人主张,它在整个人属(Homo)谱系中一以贯之。人类的偏手极强,而真正“两手都灵”的双利者非常罕见,这一点放在灵长类里,反而显得不寻常。
野外或圈养环境中,某些猴类和猿类的部分群体确实会表现出方向性的用手偏好;黑猩猩中也有有限的右手偏向证据,并且和它们的脑部不对称有所关联。但这些研究往往样本偏小,同一只猿在不同任务里(比手势、伸手取物、用工具)用手习惯还会变,甚至有人担心圈养个体会被人类饲养者“带”成右手。
一项重要的元分析得出过一个关键结论:非人类的类人猿在群体层面并不表现出方向性偏手,但在需要双手协调的任务中,个体确实会表现出很强的偏手。换句话说,“偏不偏某只手”和“群体上是不是都偏同一只手”是两码事,而人类偏侧的强度与一致性,至今没有任何灵长类能与之相提并论。它因此被称作一个未解的演化奇点。
一个是方向:偏的是左手还是右手;另一个是强度:偏得有多坚决。一个物种完全可能个个都很坚决,但有的坚决用左、有的坚决用右,最终的结果是,群体层面看上去毫无方向。人类的特殊,恰恰是这两个维度同时被拉到了极端。
答案太多,反而成了问题
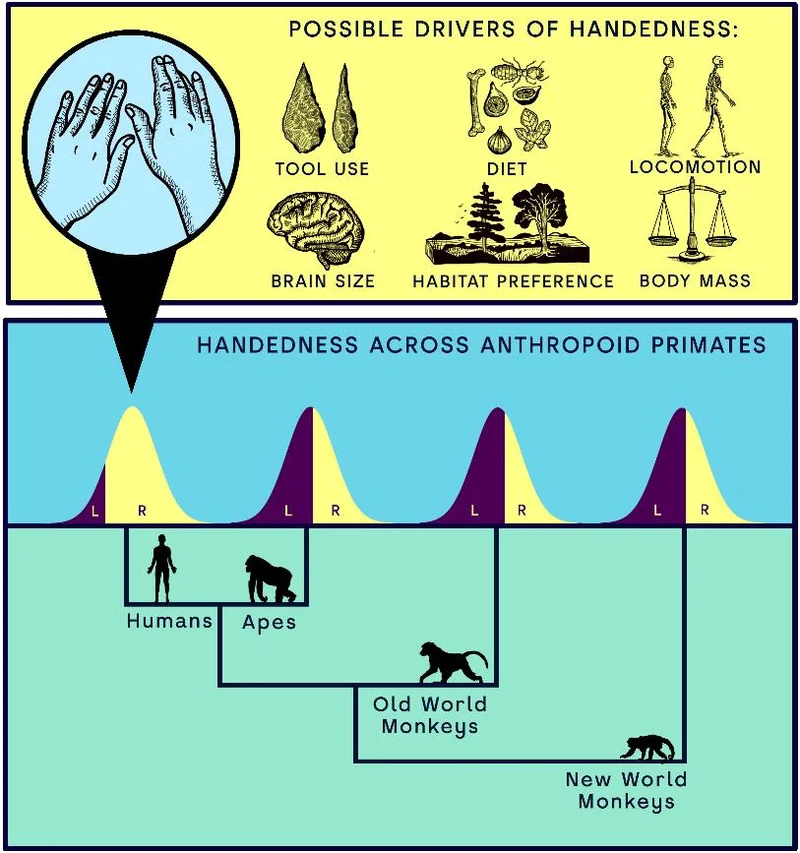
为什么是右手?有人归因于工具的制造与使用,有人归因于投掷,有人追溯到语言和手势在大脑左半球的偏侧化,有人提出抱婴假说、性内竞争假说、觅食方式假说、社会学习假说……这些说法在文献里长期并存。问题在于,其中相当一部分是以模糊的、描述性的方式提出的,很难被设计成一个可证伪的检验。最终结果就是,几十种假说彼此竞争,却谁也没能真正把对手比下去。
例如,索托(Soto)等人 2022 年的元分析显示,非人类的类人猿在群体层面并不存在方向性偏手,但个体在双手协调任务中总是表现出很强的偏手。卡斯帕(Caspar)等人 2022 年发表于 eLife 的系统发育分析则证实,人类的右手偏好是一个极端个案,群体层面的偏手在非人灵长类中很罕见;偏侧性与工具使用、栖息基质、脑容量之间没有强关联;而且,陆生物种的偏手比树栖物种更弱。
这些结论无法得到有机的统一的根源,或许在于研究方法。非人灵长类的偏手研究本就比人类少得多,尤其缺乏跨物种、长时段的系统数据;现有研究又彼此孤立。如果把这些来源不一的数据简单堆在一起进行比较,反而会引入巨大的偏差。
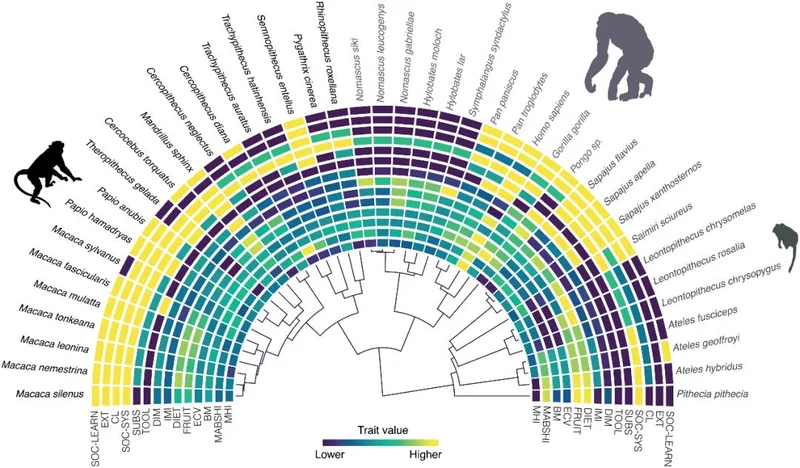
图 | 研究分析的性状及涉及类人猿物种的演化树(来源:DOI: 10.1371/journal.pbio.3003771)
在此基础上,牛津大学团队选择把这两种常用的研究工具合二为一。他们整合了上述两套最新数据,最终得到一个覆盖 41 种类人猿、2,025 个个体的标准化数据集;无论人还是猿,都只采用同一个简单、跨物种通用的测试:“握管任务”(tube task)。这要求受试者一只手握住一根透明管子,用另一只手的手指去掏出管内的食物,做“掏”这个动作的手就被记为偏好手。
人类,一个站在所有规律之外的点
实验的结果出人意料。跨越 41 个物种来看,类人猿整体上并没有统一的左右方向偏好,方向指标几乎为零。能呈现可信右偏的物种,整个数据集里只有一个,就是人类,其方向指标高达 0.76(+1 代表全员极端右利、−1 代表全员极端左利、0 代表无方向)。黑猩猩、大猩猩、戴安娜长尾猴等常见灵长类物种只有微弱到中等的右偏倾向。此外,明确呈现可信左偏的物种也只有两个:红毛猩猩和川金丝猴。
但从强度看,类人猿中绝大多数物种个体其实都有很强的偏手,只是方向各异,最终在群体层面互相抵消了。人类的强度值约 0.94,居于最高之列,但偏手倾向最强的是东爪哇叶猴,约为 0.98。在整体趋势上,研究证实了卡斯帕(Caspar)的观点:人类以及蜘蛛猴、叶猴这类偏树栖的物种偏手最强,狮尾狒、狒狒这类偏陆生的物种偏手则较弱。
当研究者用所有旧假说逐一解释这些数据时,没有任何一个假说显著优于其他,无论分析里是否包含人类。这印证了一个长期被怀疑、却很少被严格证明的判断,过去许多关于人类偏手的解释,可能都是一种循环论证:它们看上去能解释人,是因为它们本来就是为了解释人而量身定做的。
把“走路”和“脑容量”放进模型,特殊性就消失了
研究者注意到一个反复出现的现象:只要模型里同时纳入两个变量,人类的异常值身份就会发生剧烈地改变。其中一个是脑容量(用颅腔容积衡量),另一个是“肢间指数”,即前肢相对后肢的长度,肢间指数越低,腿越比胳膊长,这也是用两条腿走路的经典解剖标记。
在一个只看猿猴规律的模型里,按规律推算,人类应该和大多数灵长类一样没有方向偏好;可人类的实测值是 0.76,相差甚远。这表明,必然有一种强烈的、特有的选择压力,推动人类产生如此巨大的偏离。
然而,一旦把脑容量和肢间指数,以及社会结构放进模型,推算出的方向指标立刻跳到 0.74。强度维度同样如此,排除人类时的推算值约 0.43、人类的实测值约为 0.94,差距悬殊;而把肢间指数加进来后,推算值升到 0.86,人类就不再是离群点。
而且,这两个因素的分工并不相同。运动方式对偏手强度的解释力极强,而脑容量主要与方向绑定。
回到几百万年前,故事被分成两段
既然模型能与现存物种的偏手情况相拟合,研究者就顺势用它反推了古人类祖先。
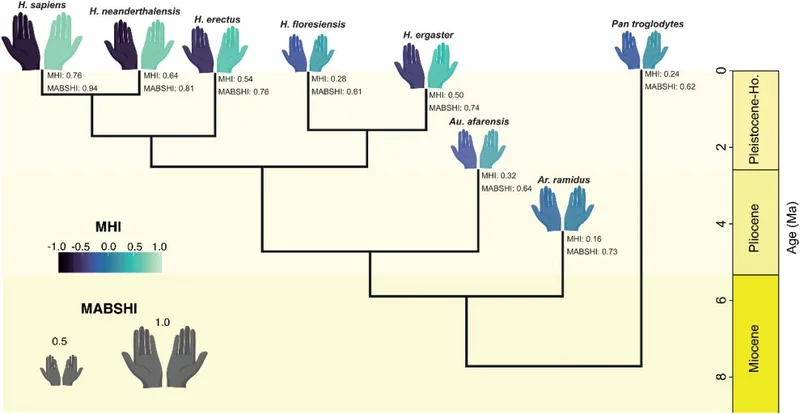
图 | 我们祖先的惯用手推演(来源:DOI: 10.1371/journal.pbio.3003771)
结果显示,几乎从人族与黑猩猩族的最近共同祖先时代起,强烈偏手的情况就稳定存在了,且一直没怎么变。从距今数百万年前的地猿开始,古人类的偏手强度推算值已经很高,在 0.7 上下,并且在之后的南方古猿阿法种、匠人、直立人、尼安德特人中保持高位。
再看方向,群体性右偏是一个更晚时期才被锁定的特征,而它和脑量扩张的节奏高度同步。早期的地猿、南方古猿只有温和的右偏,和今天的大型猿类相差不远。转折发生在人属出现、大脑显著增大之后:匠人约 0.50、直立人约 0.54、尼安德特人约 0.64,再到智人的 0.76。
这条规律还存在一个重要的反例——弗洛勒斯人。这是一类出土于印度尼西亚、身材矮小的小脑容量物种。它虽然属于人属,方向指标却只有约 0.28,明显偏低。但它的特征恰好验证了这项研究的结论:弗洛勒斯人脑容量明显更小,运动方式是直立行走与攀爬的混合类型,因此方向偏侧也不显著,模型完全解释得通。
谜底揭开后,演化的故事被分为两段。首先,当我们的祖先站起来,开始用两条腿走路,双手第一次从负责移动中解放,这创造了对精细、偏侧化手部操作的新选择压力,这也解释了为什么偏手强度升高得很早,且稳定性持续至今。
随着脑容量增大、皮层重新组织,半球特化加深,原本方向随机的强偏手演变成群体一致的右偏,这解释了为什么直到人属出现、脑子变大之后,方向偏好才陡然增强。值得一提的是,这项研究第一次直观展示了脑量扩张与偏手方向之间精确的时间对应关系。
前文有一个反直觉的发现,人类明明是彻底的陆生物种,偏手强度为何像树栖物种一样高?对此,研究人员给出的解释是,树栖生活需要一只手稳住身体、另一只手做精细动作,天然需要较强的偏手;而双足行走以另一种方式“重演”了这种状态。这也和双足行走或许最初起源于树栖场景的观点不谋而合。
最后,模型还纳入了社会结构这一变量。然而,跨文化看,右手主导依然稳定地存在,迄今没有任何一个有记载的人类社会,出现了集体左利手现象。这说明,考虑到人类是唯一拥有累积性文化演化的物种,文化虽然不是源头,但可能承担了放大器的作用,进一步放大、稳定了这种右偏。