驯化:野生苹果的召唤Robin Allaby
种群统计学、选择作用以及野生型与驯化型之间的基因流相互交织,会掩盖物种的驯化过程。一项新研究的作者揭示,现代苹果是通过与欧洲野生苹果种群发生适应性渐渗(adaptive introgression)而形成的。
驯化可被视为一条演化轨迹,植物、动物或微生物沿此轨迹适应人类生态位¹。尽管驯化物种的野生祖先种通常较为明确,但其具体的适应过程却往往难以厘清。当涉及多个野生种群与亚种时,关于适应本质的作用机制问题便会浮现,而这会直接影响驯化物种的利用价值。以葡萄为例,正是与欧洲野生种群的渐渗杂交,使其果皮更厚、在湿润气候下抗病性更强,且恰好更适合酿酒 ²。亚麻通过与野生型的渐渗杂交改变了开花时间,顺带改变了纤维结构,从而适应高纬度地区,这一过程还与纺织革命的兴起相关 ³。大麦的生态适应性在很大程度上得益于来自广阔地理范围的众多野生生态型的基因贡献⁴。这种基因流与适应的复杂性造就了嵌合基因组(mosaic genomes),只有深入了解野生祖先种群,才能厘清演化事件如何塑造出现代驯化物种。
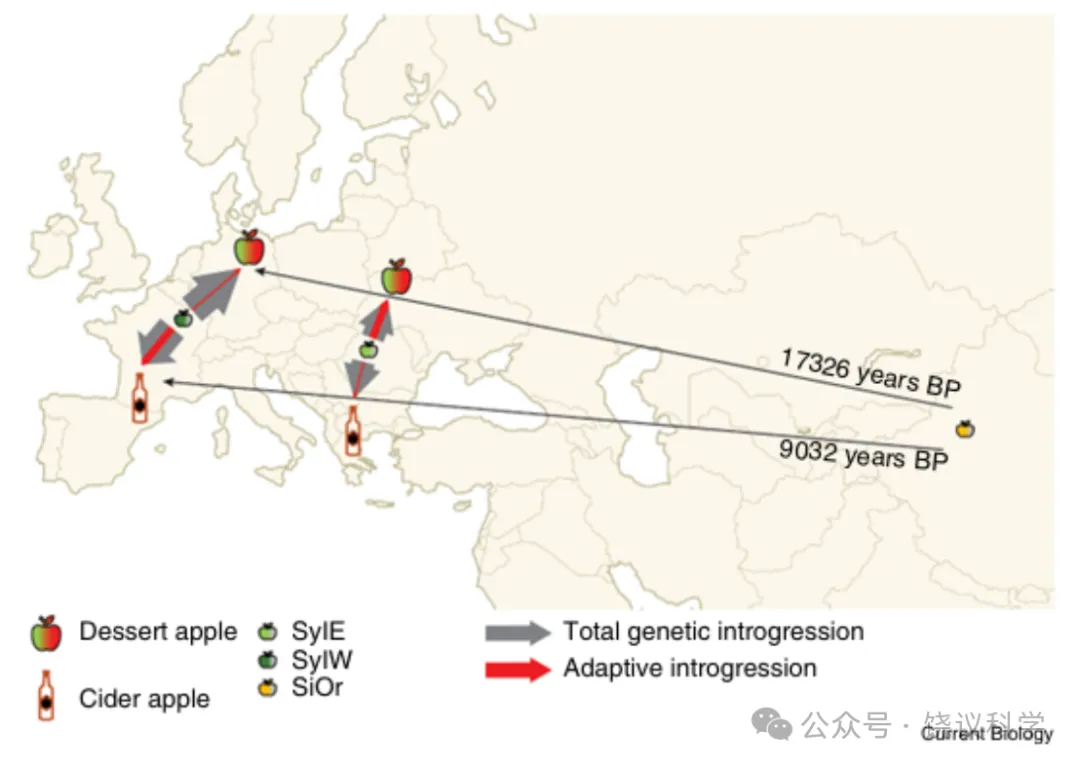
欧亚大陆的人类社会利用并广泛管理野生苹果(Malus sylvestris)已有近 8000 年历史,其也是新石器时代饮食的组成部分⁵,但栽培苹果(M. domestica)直至罗马时期才出现,晚于葡萄等已采用嫁接技术的作物⁶。欧亚大陆分布着多个苹果野生亚种,包括欧洲的M. sylvestris、高加索与中东地区的M. orientalis、中亚及中国的M. sieversii,以及中国东部的M. baccata。栽培苹果与M. sieversii的遗传相似性最高,表明其可能起源于中亚天山山脉⁷。然而,苹果的所有亚种均可互交,且栽培苹果与包括M. sylvestris在内的多个类群共享部分遗传祖先⁸,这意味着苹果在向欧洲传播的过程中有着复杂的演化历史。此外,栽培苹果可分为鲜食苹果与酿酒苹果两个混合基因库,二者有着不同的演化历程⁹。要揭开苹果起源的 “黑箱”,必须深入解析M. sylvestris对栽培苹果的遗传贡献。但迄今为止,研究的局限在于缺乏合适的M. sylvestris基因组样本。本期《Current Biology》中,一项纳入M. sylvestris的大规模基因组研究揭示,鲜食苹果与酿酒苹果代表了苹果向欧洲传播的两个时间上独立的扩散浪潮,二者从野生M. sylvestris种群中获得了不同的适应性渐渗特征¹⁰。
Chen 等 ¹⁰分析了 218 份苹果基因组,其中包含 87 份M. sylvestris基因组。研究团队鉴定出 6 个独立种群。野生亚种M. baccata是其他亚种的独立外类群。鲜食苹果与酿酒苹果的基因组分别形成两个独立但存在混合的基因库(分别为 DomD 与 DomC),这与此前研究结果一致。M. sieversii与M. orientalis归为一个未分化的单一种群(SiOR),而M. sylvestris则形成欧洲西部(SylW)与欧洲东部(SylE)两个独立基因库,二者约在 1.26 万年前的冰期后扩张中分化形成。鲜食苹果与酿酒苹果无法从祖先 SiOR 种群中区分开来,表明二者可能代表不同时期的两次独立扩散。鲜食苹果的演化支系率先分化,约在 1.73 万年前的晚冰期;酿酒苹果支系则在 9000 年前的全新世较晚时期分化。因此,原始栽培苹果种群在向西迁徙的过程中,历经了由冰期气候循环塑造的两种截然不同的环境。在此研究中,生态位建模可有效揭示每次扩散所面临的选择压力,该方法已成功应用于不同水稻类型的研究 ¹¹。
首先,苹果在迁徙过程中进入已有本地适应种群定居的新环境,发生适应性渐渗并不意外;其次,鲜食苹果与酿酒苹果种群的适应重点存在差异,也合乎情理。鲜食苹果与酿酒苹果种群均接收了来自 SylE 与 SylW 的渐渗信号,且二者从 SylW 获得的渐渗比例均显著更高 ¹⁰。两个驯化类群接收的总渐渗量差异微小,但涉及的野生祖先基因组片段不同。然而,在考虑选择作用时,二者呈现出显著差异。其一,鲜食苹果的早期扩散与硬选择清除相关,即对稀有或新突变的强选择;相比之下,酿酒苹果的后期扩散更多与软选择清除相关,即对已处于中等频率的变异的选择。这种差异可能反映出,首次扩散历经晚冰期更为极端的气候条件,而后期扩散则处于全新世早期更温和的气候环境。值得注意的是,鲜食苹果早期向欧洲扩散时,与渐渗相关的选择清除主要来自其最先接触的野生种群 SylE;相反,后期扩散的酿酒苹果主要从更西侧的 SylW 获得适应性渐渗,且整体适应性渐渗区域更少。这些适应性渐渗大多能帮助原始栽培种群在被引入的新环境中存活,涉及抗病性、耐温性与开花时间等性状,而这些适应不可避免地影响了苹果的利用性状。在鲜食苹果的 23 个渐渗区域中,研究团队重点指出 16 号染色体上包含酸度标记Ma的区域;在酿酒苹果的 14 个渐渗区域中,17 号染色体上存在一个与果实质地相关的基因座 ¹⁰。这些来自冰期不同阶段、针对不同环境的差异化适应,是否偶然推动苹果分别演化成适合鲜食的甜味类型与适合酿酒的酸味类型,仍是一个待解的问题。
不同种群间因能为彼此提供环境适应性而产生的基因交流,也可能由基因组修复的共同需求驱动。种群无论因选择还是其他因素发生瓶颈效应时,有害变异都会在种群中固定,进而增加突变负荷¹²,降低种群适合度 ¹³。两个具有独立突变负荷积累历史的种群,具备相互拯救的潜力。来自高粱亚群的证据表明,基因流保留的特征偏向于突变负荷的修复,这提示种群间存在驱动基因交流的吸引力 ¹⁴。Chen 等 ¹⁰的研究发现,鲜食苹果与酿酒苹果种群的遗传多样性均高于野生种群,这可能反映了种群间的杂交;同时二者的突变负荷更低,这与驯化物种的普遍预期相悖。该研究尚未验证栽培苹果中来自祖先 SiOR 的突变负荷,是否被 SylW 与 SylE 的基因流所抵消。但颇具浪漫色彩的是,驯化苹果在穿越欧洲的旅程中,或许响应了 “荒野的召唤”,找回了基因组中缺失或受损的部分 ¹⁵。
图 1 栽培苹果向欧洲的传播路径
栽培苹果的两次扩散均起源于塞威士苹果(SiOR),且发生于不同时期,最终分别形成鲜食苹果与酿酒苹果。在欧洲,二者从欧洲东部野苹果(SylE)与欧洲西部野苹果(SylW)种群中发生了模式迥异的适应性渐渗。(地图来源:Ultimaps.com)
REFERENCES......